2.3
Potenciales de transferencia de fosfato
Para comparar el potencial de transferencia de
fosforilos de los compuestos fosforilados se utilizan las energías libres estándar
de hidrólisis. Al realizar estas comparaciones se observa que el ATP no es el único
compuesto con un alto potencial de transferencia de fosforilos. De hecho,
algunos compuestos de los seres vivos poseen un potencial de transferencia
mayor que el ATP. Algunos de esto compuestos son el fosfoenolpiruvato (PEP),
1,3-bisfosfoglicerato y la creatina fosfato. Así, el PEP puede transferir un
grupo fosforilo al ADP para formar ATP. Esta es una de las formas de producir
ATP durante la degradación de los azucares. Resulta significativo que el ATP
tenga un potencial de transferencia de fosforilos que es intermedio entre el de
las moléculas biológicamente importantes. Esta posición intermedia capacita al
ATP para funcionar eficientemente como transportador de grupos fosforilo.
La cantidad de ATP fosforilo disponible en el
músculo asegura su capacidad de contracción durante menos de un segundo. La creatina
fosfato en el musculo de los vertebrados es un reservorio de grupos fosforilo
de alta energía que puede transferirse fácilmente al ATP. Cada vez que
realizamos un ejercicio intenso utilizamos la creatina fosfato para regenerar el ATP a partir del ADP. La enzima
creatina quinasa cataliza esta reacción:

Además del ATP existen
otros compuestos con potenciales de transferencia de grupos fosfato significativamente
mayores que el ATP, pero poseen influencias desestabilizadoras adicionales como
son:
1.
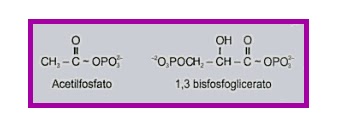
Acilfosfatos
El ácido
1,3-bisfosfoglicérico es un ejemplo de un acilfosfato. Su energía libre estándar
de hidrólisis es de -11.8 kcal/mol. La tensión de enlace en el acil fosfato es
un factor importante que contribuye a la gran energía libre estándar negativa
de hidrólisis de este grupo de compuestos. Puede considerarse que elenlace C=O
del grupo acil fosfato tiene un carácter polar considerable, debido a la
tendencia de los electrones del doble enlace
a aproximarse al oxígeno electronegativo.
2.
Enolfosfatos
El elevado
potencial de transferencia del grupo fosfato
de los enolfosfatos, como el fosfoenolpiruvato, deriva de que el enol,
producto de su hidrólisis, es menos estable que su tautómero ceto.
3.
Fosfoguanidinas
Los elevados
potenciales de transferencia de fosfato de las fosfoguanidinas, como la fosfocreatina y la fosfoarginina, son el
resultado, en gran parte, de la competencia de resonancia en sus grupos
guanidino, que son aún mas pronunciadas que en el grupo fosfato de los
fosfoanhídridos.
Son 3 las
funciones principales de –P que toman parte en la conservación o captura de la energía:
1.
Fosforilación oxidativa
A pesar de que
fue mencionada anteriormente en la formación de ATP, cabe señalar que es la
fuente cuantitativa mayor de –P en los organismos aerobios. La energía libre
para conducir este proceso procede de la oxidación de la cadena respiratoria
dentro de las mitocondrias.
2.
Glucólisis
Hay una formación
neta de dos –P como resultado de la formación de lactato a partir de una molécula
de glucosa, generadas en 2 reacciones catalizadas por la fosfoglicerato cinasa
y la piruvatocinasa, respectivamente.
3.
Ciclo de Krebs
En el ciclo se
genera un –P directamente en el paso catalizado por la succiniltiocinasa.

Referencias:
Libro: bioquímica, pág. 416, disponible
en:
libro: bioquímica de los procesos
metabólicos, págs.. 24-25, disponible en:
No hay comentarios:
Publicar un comentario