6.1 Conceptos de permeabilidad y potencial de membrana

Permeabilidad
La permeabilidad selectiva de las
membranas determina qué tipo de sustancias pueden entrar y salir de la célula.
El ingreso de sustancias necesarias, el pasaje de agua y la salida de los
productos de desecho, se verán posibilitados y regulados por medio de la membrana
plasmática.
Los distintos tipos de pasaje a través de
la membrana pueden clasificarse en dos grandes grupos. En uno de ellos, los
iones o moléculas pequeñas son transportados por las membranas, sin que estas
experimenten deformaciones evidentes. En el otro mecanismo, denominado
transporte en masa, las macromoléculas o partículas de mayor tamaño son
incorporadas a la célula o eliminadas de ella por medio de procesos que incluyen
cambios visibles en las membranas.
La membrana plasmática regula la entrada
y salida de materiales, permitiendo la entrada de unos y restringiendo el paso
de otros. Esta propiedad se llama permeabilidad selectiva.

La membrana es permeable cuando permite
el paso, más o menos fácil, de una sustancia. La permeabilidad de la membrana
depende de varios factores relacionados con las propiedades físico-químicas de
la sustancia:
Solubilidad en los lípidos: Las
sustancias que se disuelven en los lípidos (moléculas hidrófobas, no polares)
penetran con facilidad en la membrana dado que esta está compuesta en su mayor
parte por fosfolípidos.
Tamaño: la mayor parte de las moléculas
de gran tamaño no pasan a través de la membrana. Sólo un pequeño número de moléculas
no polares de pequeño tamaño pueden atravesar la capa de fosfolípidos
Carga: Las moléculas cargadas y los iones
no pueden pasar, en condiciones normales, a través de la membrana. Sin embargo,
algunas sustancias cargadas pueden pasar por los canales proteicos o con la
ayuda de una proteína transportadora.
También depende la permeabilidad de una
membrana de la naturaleza de las proteínas de membrana existentes

Canales: algunas proteínas forman canales
llenos de agua por donde pueden pasar sustancias polares o cargadas
eléctricamente que no atraviesan la capa de fosfolípidos.

Transportadoras: otras proteínas se unen
a la sustancia de un lado de la membrana y la llevan del otro lado donde la
liberan.
En general, estos canales y proteínas
transportadoras muy altamente selectivas permitiendo el paso a una única
sustancia.
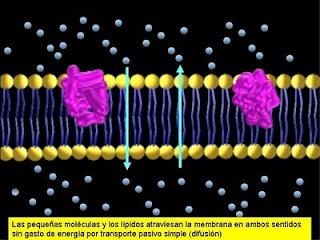
Permeabilidad de moléculas pequeñas
El pasaje de estas moléculas va a
depender de si las mismas se movilizan a favor o en contra de su gradiente de
concentración. Cuando lo hagan a favor del gradiente, pasarán en forma pasiva, es decir que no habrá gasto de energía. Si, por el
contrario, las moléculas se movilizan en contra del gradiente de
concentración será necesario el aporte
de energía y el pasaje será activo.
Potencial de membrana
El
potencial de membrana hace referencia a la diferencia de cargas eléctricas a
través de la membrana plasmática. Fuera de la membrana esta cargado
positivamente debido a los iones Na+ y Ca2+ en grandes concentraciones y por
dentro de la membrana es negativo debido a grupos cargados negativamente en el
citoplasma.
El potencial de membrana o de reposo
depende de:
a) La redistribución de los iones a
través de la membrana. El Na+ y Ca+2 son más abundantes afuera. El K+, PO4-,
SO4- y Cl- es más abundante adentro.
b) la acción de la bomba Na-K ATPasa. La
bomba tira 3 sodios hacia afuera y 2 potasios hacia adentro simultáneamente,
por eso se carga positivo afuera.

c) los canales de K siempre abiertos. El
potasio sale por los canales abiertos atraído por el gradiente químico y luego
incrementa la repulsión eléctrica con los iones de sodio, incrementándose el
gradiente eléctrico que repele al potasio. Se crea un equilibrio entre ambos
gradientes y el potasio deja de salir quedándose más concentrado en el interior
de la célula.
El potencial es de alrededor de -75 mV en
las neuronas y es solo unas 20 veces menor que una pila de 1,5 V. Todas las células
vivas tienen un potencial de reposo característico.

La electroneutralidad
La existencia de una diferencia de
potencial es ante todo una indicación de la existencia de una separación de
cargas. De alguna manera las membranas biológicas contribuyen a que se mantenga
un exceso relativo de cargas negativas en el interior celular con respecto al
medio extracelular. Sin embargo, es un hecho empírico muchas veces elevado a
nivel de principio, el que cualquier porción macroscópica de una solución es
electroneutra. La contradicción aparente se despeja cuando se considera lo que
se entiende por “macroscópica” en la frase precedente. El entorno inmediato de
las membranas biológicas puede ser lo suficientemente pequeño para ser
considerado no macroscópico, y sin embargo soportar una diferencia de potencial
medible con los instrumentos usuales. Un modesto (y muy aproximado) cálculo
aclarará las cosas. Ver para ello el ejercicio 1. El resultado es que con
variaciones pequeñas de la concentración es posible mantener una diferencia de
potencial del orden de 100 mV . El pequeño exceso de carga se distribuye
rápidamente en una delgada capa cercana a las proximidades de la membrana
plasmática, haciendo al interior celular electroneutro.
Referencias:
No hay comentarios:
Publicar un comentario